Lipit metabolizması ve oksidasyonu
Lipitlerde oksidasyon
Lipitler lezzet, tokluk ve beslenme katkısı olarak gıda maddeleri arasında önemli bir yere sahiptir. Lipit oksidasyonu gıda endüstrisinin pek çok sektöründe önemli bir sorundur. Lipit oksidasyonu terimi aktif oksijen türleriyle lipitlerde doymamış yağ açil grupları arasındaki kimyasal interaksiyonların kompleks zincirini tanımlamak için kullanılır (NAWAR, 1996; McCLEMENTS ve DECKER, 2000; MİN ve BOFF, 2002).
Arzu edilen besinsel ve fiziksel özelliklere sahip ürünlerin geliştirilmesi için lipit oksidasyon mekanizmasının iyi bilinmesi gerekir. Yağlarda meydana gelen oksidatif tepkimeler; otooksidasyon, termal oksidasyon, fotooksidasyon ve enzimatik oksidasyon olmak üzere dört şekilde meydana gelmektedir (SU, 2003). Otoksidasyon, kendiliğinden oluşan bir reaksiyon olup, ortamdaki ışık, ısı ve çok değerlikli metaller tarafından katalizlenmekte ve moleküler oksijenin doymamış yağ asitlerine katılımı sonucu, hidroperoksitlerin oluşumunu kapsamaktadır. Otooksidasyon, aslında bir serbest radikal zincir reaksiyonudur ve üç aşamada gerçekleşmektedir. Bu aşamalar başlangıç, gelişme ve sonlanma basamakları şeklindedir (FRANKEL, 1998; KAMAL-ELDİN ve POKORNY, 2005).
Başlangıç aşamasında serbest radikaller yağda çözünmüş olan moleküler oksijeni yakalayarak peroksi radikallerini oluşturmaktadır (ROO*). Gelişme aşamasında peroksi radikali doymamış yağ asitlerinden bir hidrojen kopararak bir hidroperoksit ile bir serbest radikal oluşturmaktadır. Bu basamakta gerçekleşen net reaksiyon oksijen ile doyamamış yağ asitlerinin tepkimeye girmesini ve oksidasyonunun birincil ürünleri olan hidroperoksitlerin oluşmasını kapsamaktadır. Eğer ortamdaki serbest radikaller birbirleri ile tepkimeye girerek reaktif olmayan ürünler meydana getirirlerse, bitiş aşaması başlamış demektir. Ayrıca, oluşan oksidasyon ürünleri reaksiyonu katalize etmektedir.
Lipit oksidasyonuna neden olan başlıca etkenler
1. Su aktivitesi
Gıda maddeleri sahip oldukları nem düzeylerine göre yüksek, orta ve düşük nemli gıdalar olarak sınıflandırılmaktadırlar. Yüksek nemli gıdalarda (aw>0,85) mikrobiyal aktivite fazla olmasından dolayı lipit oksidasyonu önemli sorun oluşturmamaktadır. Orta nemli gıdalarda (0,65<aw<0,85) lipit oksidasyon hızı fazladır. Genel olarak düşük nem içeren gıdalarda aw<0.1’den düşük olduğu zaman lipit oksidasyon hızı yüksektir. 0,1-1,2 aw’lerde suyun artmasıyla dokudaki oksijen çıkmakta, reaksiyonu katalizleyen metaller hidrate olmakta ve yağ-su ara yüzeyinde oluşan hidroperoksitlere hidrojen bağlanarak hidroperoksit sayısında azalma olmaktadır (LABUZA, 1982). 0.2-0.6 arasındaki aw’lerde metaller, şelat yapan ajanlar ve antioksidantlar suda çözündüğünden metallerin bağlandığı ve antioksidantların etkin hale geçerek lipit oksidasyonunu yavaşlatmaktadır. Aw değeri 0,6’nın üzerine çıktığı zaman oksidasyon hızı tekrar artma eğilimi gösterir. Bunun nedeni makromoleküllerin şişerek peroksitlerin birleşebileceği katalitik yüzeyler oluşturması ve giderek artan su miktarıyla kataliz mobilitesini arttırması olarak belirtilmiştir (KAREL ve YONG, 1981; LEUNG, 1987).
2.Sıcaklık
Sıcaklığın etkisiyle, alkil peroksitlerinin parçalanmalarında açığa çıkan serbest yağ asitleri, trigliseritlere göre reaksiyonu daha çok hızlandırarak biyokatalizör görevi görürler. Sıcaklığın bir başka fonksiyonu, konsantrasyona bağlı olarak proksidant veya antioksidant etki yapabilen suyu uçurmasıdır (ANGELO ve ark., 1975a). Sabit sıcaklıkta yapılan bir çalışmada, nem oranı yüksek olan örneklerin serbest yağ asitleri değerleri daha yüksek bulunur (ROBERTSON ve ark., 1984). Nem oranının sabit tutulduğu çalışmalarda ise en yüksek sıcaklıkta depolanan örneklerin en yüksek serbest yağ asitleri değerine sahip olduğu görülmüştür (HUNTER ve ark., 1951).
3.Metaller
Metaller proses aşamasında bulaşabileceği gibi gıdanın içinde de doğal olarak bulunabilmektedir. Geçiş metalleri, sıcaklığın etkisinden farklı olarak hidroperoksitlerin bozulması ile oluşan ikincil ürünlerin oluşumunu katalize etmelerinden dolayı sekonder katalizör olarak adlandırılırlar (ÜNAL ve NERGİS, 1986). Metal iyonları substrattan bir elektron alarak veya vererek zincir reaksiyonlarını başlatmaktadırlar ya da serbest radikal oluşturmaktadırlar. Serbest metaller suyun dilüsyon etkisine duyarlıdırlar. Düşük su aktivitelerinde metal hidratasyonu nedeniyle antioksidan etki söz konusudur.
4.Enzimler
Lipit oksidasyonunu katalizleyen enzimlerin başında lipoksidaz ve peroksidaz gelmektedir (PATTEE ve ark., 1982).
Yağ Asitlerinin Katabolizması
-Uzun zincirli yağ asitlerinin asetil CoA’ya yıkılmaları pek çok organizma için enerji metabolizmasının en önemli kısmını oluşturur.
-Memeli hayvanlarda kalp ve karaciğerin enerji ihtiyacının %80’i yağ asitlerinin yıkımı ile karşılanır.
-Yağ asitlerinin oksidasyonu sırasında açığa çıkan elektronlar solunum zincirinden geçerek ATP sentezini sağlar.
-Yağ asitlerinden sentezlenen asetil CoA sitrik asit döngüsünde tamamen karbondioksite oksitlenerek enerji üretimi yoluna katkıda bulunur.
-Bazı türlerde ve dokularda Asetil CoA farklı metabolik yollara girebilir.
-Karaciğerde keton cisimciklerine çevrilebilir. Keton cisimcikleri suda çözünebilir ve glukozun olmadığı durumlarda beyin ve benzeri dokularda enerji kaynağı olarak kullanılabilir.
-Yüksek yapılı bitkilerde asetil CoA biyosentetik öncül madde olarak davranır. Yakıt olarak kullanımı ancak ikincil önemdedir.
-Her ne kadar yağ asidi oksidasyonunun rolü organizmadan organizmaya değişiklik gösterse de mekanizma neredeyse aynıdır.
-Triaçilgliseroller yüksek derecede indirgenmiş oldukları, kolay bir arada paketlenebildikleri, suda çözünmedikleri için yan reaksiyonlar vermedikleri ve depolanmaları için suya ihtiyaç duymadıkları için ideal yakıt depo molekülleridir.
-Ancak bu özellikleri yakıt olarak kullanılmaları sırasında bir takım sorunlar oluşturur.
-Triaçilgliserollerin metabolize olabilmeleri için emülsifiye olmaları gerekir.
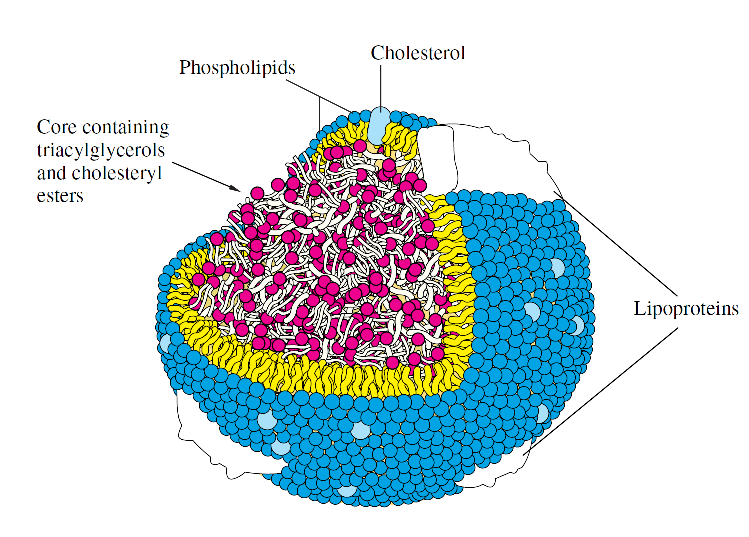
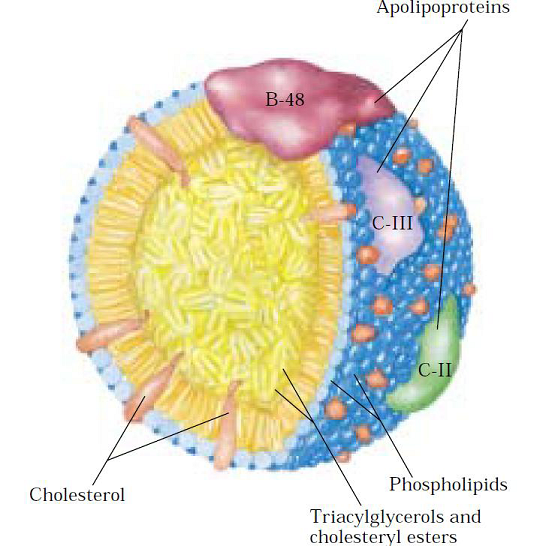
-Bağırsaklardan emilen veya depo dokularından serbest kalan yağlar kanda çözünmedikleri için proteinlere bağlı olarak taşınmak zorundadırlar.
-C-C arasındaki bağlarının oldukça kararlı olması nedeniyle C-1 karbonundan CoA’ya bağlı olarak metabolize olabilirler.
YAĞLARIN SİNDİRİLMESİ VE TAŞINMASI
-Hücreler yağ asitlerini üç kaynaktan elde edebilirler; diyetle alınan, depolardan serbest kalan veya hücre içinde üretilen yağlardan.
-Triaçilgliseroller karaciğer, kalp ve dinlenme halindeki iskelet kasları başta olmak üzere bazı organların enerji ihtiyacının yarıdan fazlasını karşılarlar.
-Depolanmış triaçilgliseroller kış uykusuna yatan hayvanların ve göç eden kuşların tek enerji kaynağıdır.
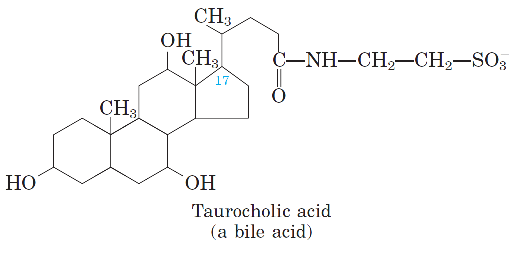
-Omurgalılarda yağların çözünürleştirilmeleri taurokolik asit gibi bazı safra tuzları sayesinde gerçekleştirilir.
-Nötral lipitler adipozitlerde (ve daha düşük miktarlarda adrenal korteksin, over ve testislerin steroit sentezleyen hücrelerinde) damlacıklar halinde depolanırlar.
-Bu damlacıkların yüzeyi perilipinler ile kaplanmıştır ve bu proteinler lipit damlasına erişimi kısıtlayarak gerekli olmayan durumlarda lipit mobilizasyonunu engellerler.
-Hormonlar metabolik enerji ihtiyacı sinyalini verdiklerinde adipoz dokudaki yağlar mobilize olur ve okside olmak üzere iskelet kası, kalp ve renal korteks gibi dokulara transfer olurlar.
-Yağ asidi oksidasyonun enzimleri hayvan hücresinde mitokondri matriksinde bulunmaktadır.
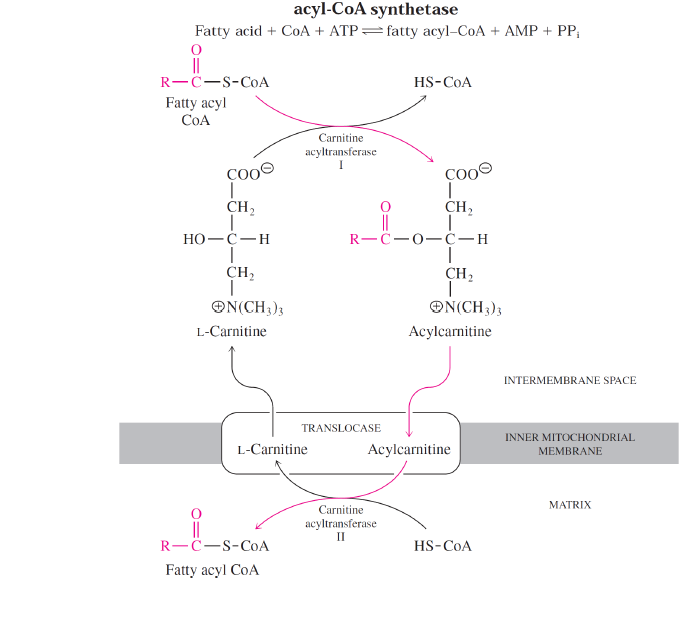
-12 veya daha az karbon sayısına sahip yağ asitleri mitokondriye taşıyıcıların yardımı olmadan girebilirler.
-14 veya daha fazla karbon sayılı olanlar ise karnitin mekik sistemini kullanrak mitokondriye girebilirler.
-Karnitin mekiği sitozol ve mitokondrideki CoA havuzlarını birbirinden ayırır.
-Mitokondrideki CoA havuzu büyük ölçüde piruvatın, yağ asitlerinin ve bazı amino asitlerin oksidatif degradasyonu için kullanılırken, sitozolik CoA havuzu yağ asidi biyosentezi için kullanılır.
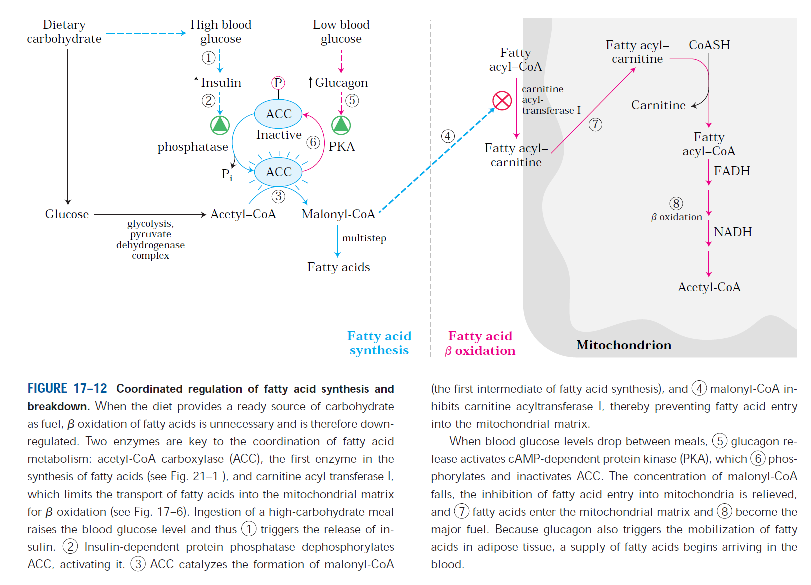
-Karnitin bağlı mitıokondri içine giriş işlemi yağ asidi oksidasyonunda hızı belirleyen basamaktır ve bu sebeple bir kontrol noktasıdır.
Yağ asitlerinin oksidasyonu
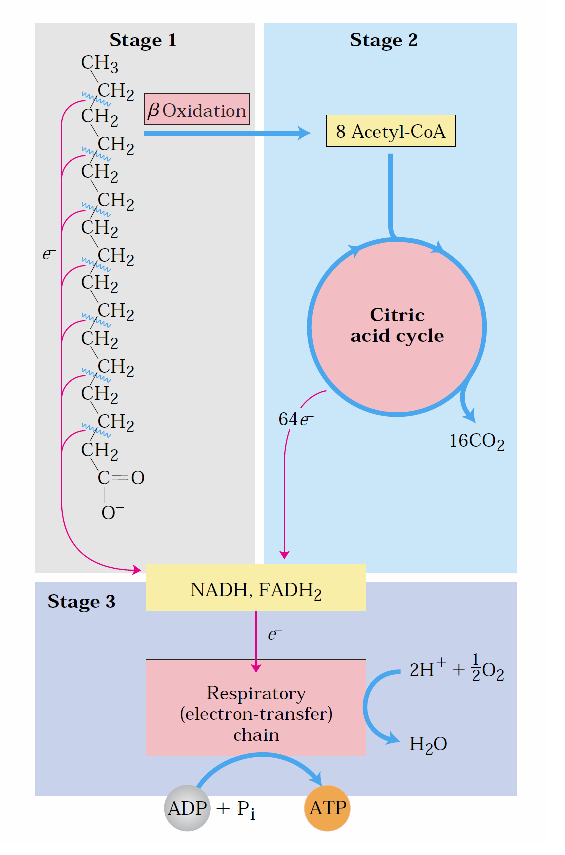
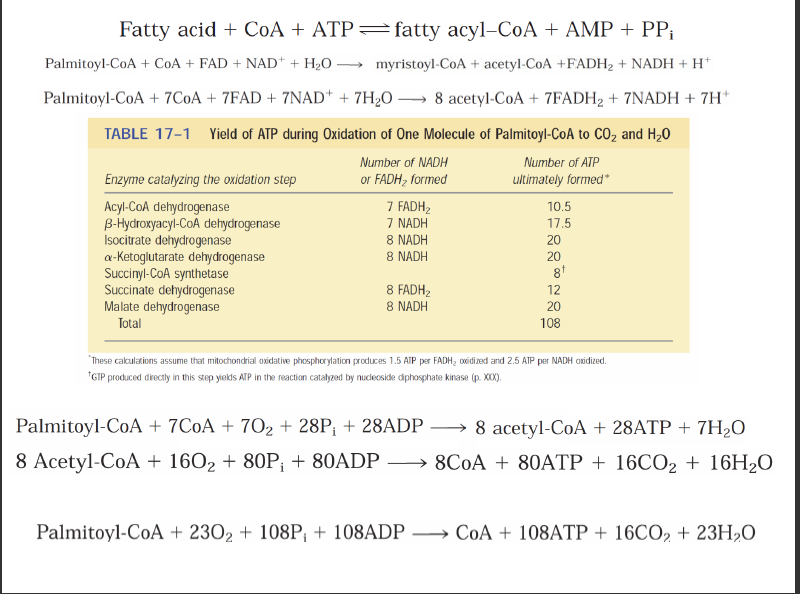
-Yağ asitlerinin mitokondriyel oksidasyonu üç basamakta gerçekleşir.
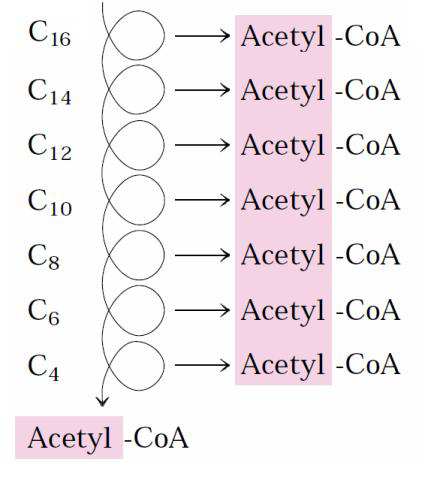
-İlk aşamada, (β oksidasyon) yağ asitlerinden iki karbonlu birimler oksidatif olarak koparılarak asetil CoA sentezlenir.
-İkinci aşamada oluşan asetil CoA’ların asetil asetil grupları sitrik asit döngüsünde karbondiokside oksitlenir.
-Üçüncü ve son aşamada ise ilk iki aşamada üretilen indirgenmiş elektron taşıyıcıları NADH ve FADH2 elektronlarını mitokondriyel solunum zincirine aktararak ATP üretimin sağlar.
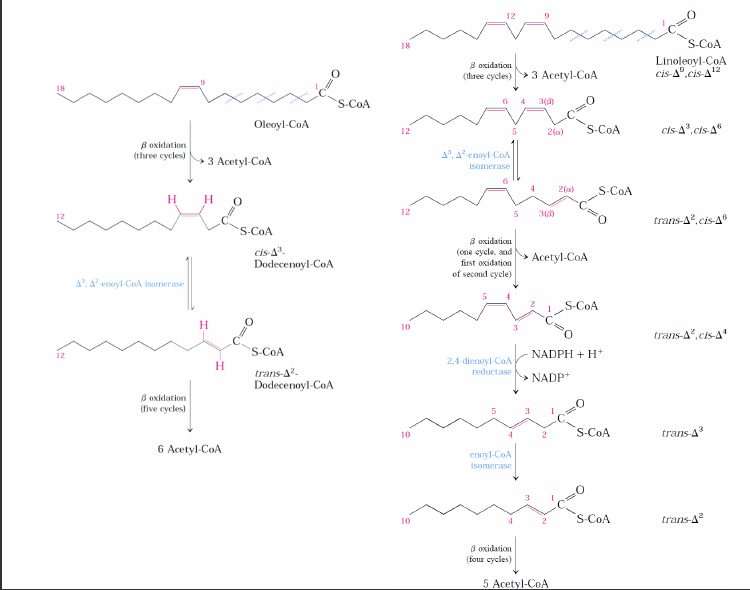
Doymamış yağ asitlerinin oksidasyonu
-Bitki ve hayvanlardaki yağları oluşturan yağ asitleri 1 veya iki doymamışlık içerirler.
-Bu bağlar cis- konfigürasyonundadır ve enoil-CoA hidrataz enzimi bu bağlara etki edemez.
-Bu engelin aşılması için biri izomeraz biri redüktaz olmak üzere iki yardımcı enzim gereklidir.
-Her ne kadar doğal olarak bulunan lipitlerin büyük çoğunluğu çift karbon sayılıysa da pek çok bitki ve deniz canlısı tek karbon sayılı yağ asitleri de içerirler.
-Geviş getiren hayvanların rümenlerinde fermentasyon sırasında yüksek miktarda propionat (3 karbonlu yağ asidi) oluşur.
-Ayrıca düşük miktarlarda propionat küflenmeyi engellemek için bazı ekmek ve tahıllara da eklenir ve dolayısıyla insan beslenmesine girer.
-Uzun zincirli tek karbon numaralı yağ asitleri tıpkı çift karbonlular gibi oksidasyona uğrar ancak en son aşamada asetil CoA yerine propionil CoA oluşur.
-Oluşan propionil CoA sitrik asit siklusuna giremez, başka bir metabolik yolla okside olur.
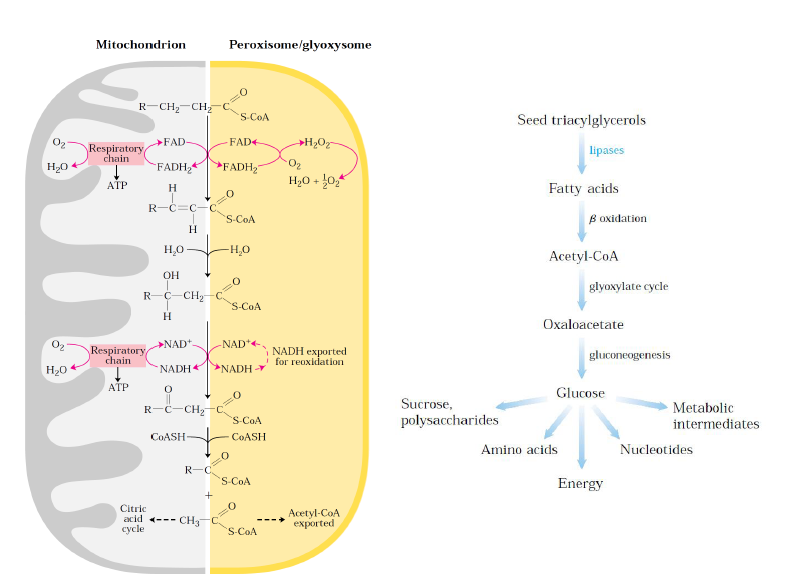
-Hayvan hücrelerinde beta oksidasyon mitokondride gerçekleşirken bitki hücrelerinde bu işlem peroksizom ve glioksozomlarda gerçekleşir.
-Peroksizomlarda ilk basamak mitokondrideki metabolik yoldan farklı gerçekleşir ve açığa çıkan enerji ATP olarak depolanmaz ancak ısı olarak dışarıya verilir.
-Bu organanellerde beta oksidasyonunun önemi enerji eldesi değil çeşitli öncül moleküllerin biyosentezidir.
-Tohumun filizlenmesi sırasında yağ asidi oksidasyonundan açığa çıkan asetil CoA glukoz, sükroz ve çeşitli metabolitlerin sentezi için kullanılır.
Yağ asitlerinin Oksidasyonu
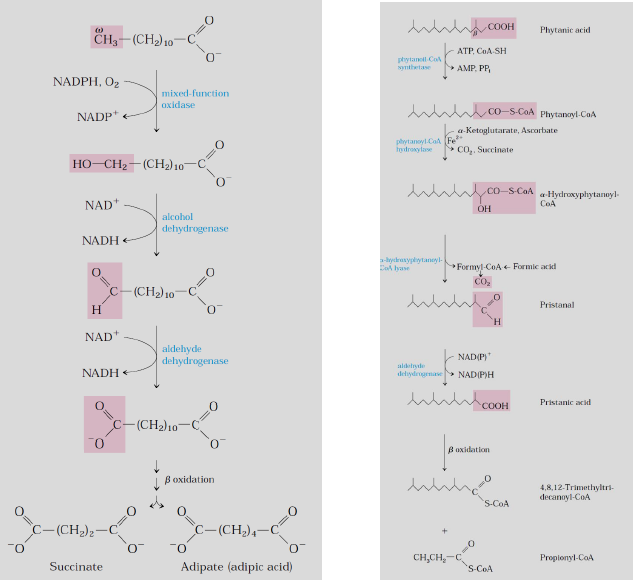
-Her ne kadar mitokondriyel β oksidasyon hayvan hücrelerinde yağ asitlerin en öenmli katabolik sonunu oluştursa da karbon zincirinin en sonundaki ω (omega) karbondan da oksidasyon gerçekleşebilir.
-Omurgalılarda ω oksidasyonun enzimleri karaciğer ve böbrek hücrelerinin endoplazmik retikulumlarında yerleşmiştir. Ve 10-12 karbonlu yağ asitlerinin oksişdasyonunu tercih ederler.
-Memelilerde ω oksidasyon son derece küçük bir rol oynar ancak β oksidasyonda sorun varsa (karnitin yetmezliği gibi) önemli bir hale gelir.
Keton Cisimcikleri
-Pek çok memeli hayvanda ve insanda karaciğerde yağ asitlerinin beta oksidasyonu ile oluşan asetil CoA ya sitrik asit siklusuna girer ya da keton cisimcikleri olarak adlandırılan aseton, asetoasetat ve D-β- hidroksibütirata çevrilerek başka dokulara gönderilir.
-Aseton solunum ile organizmadan atılabilir. Asetoasetat ve D-β- hidroksibütirat ise taşındıkları dokuda yeniden asetil CoA’ya çevrilerek enerji üretiminde kullanılırlar.
-Beyin normalde glukozu yakıt olarak tercih eder ancak açlık durumunda keton cisimciklerini de kullanmaya adapte olabilir.
Yağ Asitlerinin Biyosentezi
-Yağ asidi sentezi yağ asitleri yıkılmasının tersi değildir. Sentez ve yıkımı farklı yollarda farklı enzimler tarafından katalizlenir.
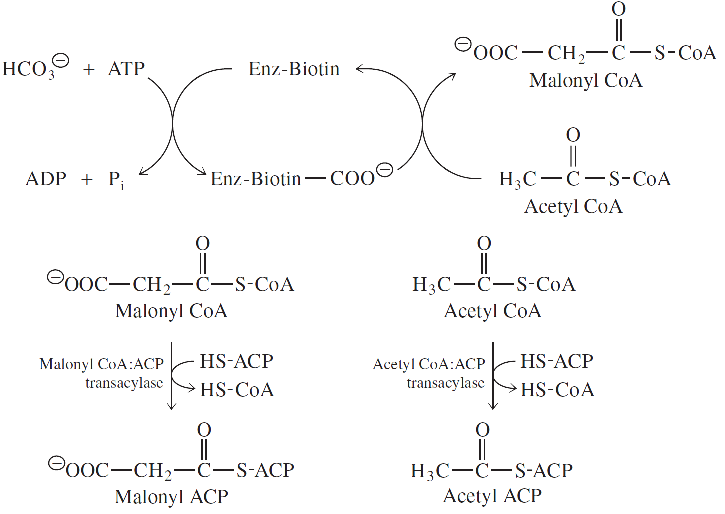
-Malonil CoA yağ asitlerin yıkımında değil sadece biyosentezinde yer alır.
-Malonil CoA’nın asetil CoA’dan tek yönlü oluşumu asetil CoA karboksilazla katalizlenir.
-Yağ asitlerinin uzun karbon zincirleri dört basamaklı bir işlemin birkaç kez çalışmasıyla oluşur. Bu tepkime dizisinde oluşturulan doymuş açil grupları aktifleştirilmiş sonraki malonil grubuyla birleşecek substrattır.
-Yağ asitlerinin sentezinde her bir döngüde yağ asidi zinciri iki karbon uzar. Ürün, zincir uzunluğu 16 karbona (palmitat) ulaştığında döngüyü terk eder.
-Asetil grubunun metil ve karboksil karbon atomları palmitatın sırasıyla C-16 ve C-15’i olur, diğer karbon atomları malonil CoA yoluyla asetil CoA’dan gelir.
-Sentez işlemlerindeki tepkimeler bir çoklu enzim kompleksi olan yağ asidi sentaz enzimi sayesinde katalizlenir.
Yağ Asidi Sentaz Asetil ve Malonil Gruplarını Alır
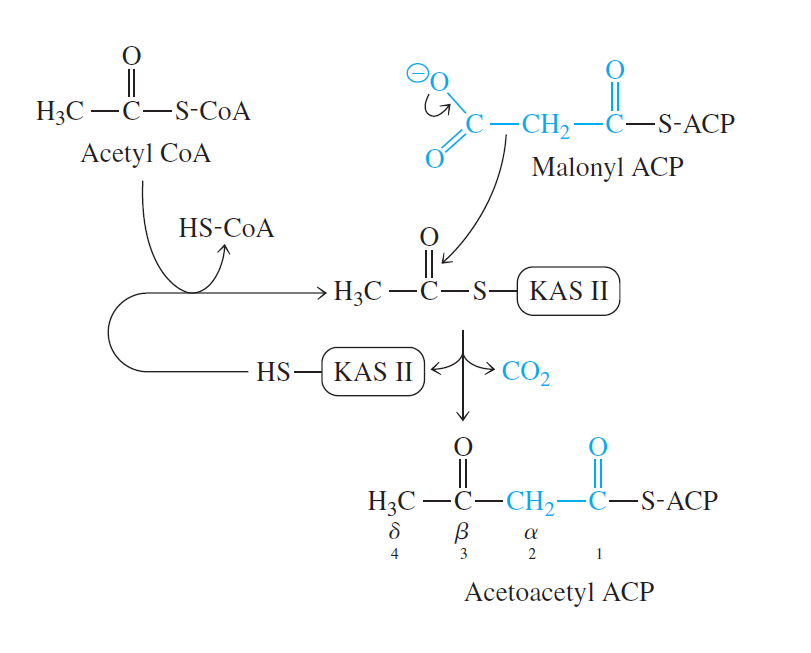
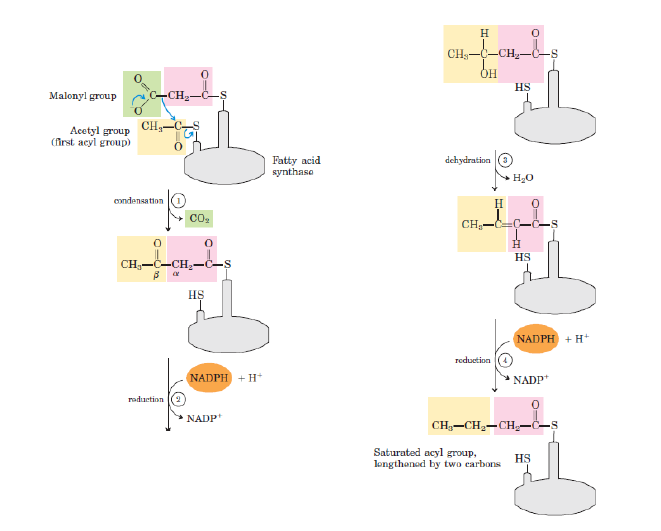
-Basamak 1: Birleştirme (Kondensasyon): Yağ asidi zincirinin oluşmasında birinci adım aktif asetil ve malonil grupların asetoasetil-ACP oluşturmak üzere birleşmesidir. Bu arada bir molekül CO2 oluşur. Bu reaksiyon β-ketoaçil-ACP sentaz (KAS) tarafından katalizlenir.
- Yağ asidi sentezini işler duruma sokmak için gerekli ekstra enerji asetil CoA ve HCO3-’dan malonil CoA sentezlemek için kullanılan ATP tarafından sağlanmaktadır.
-Basamak 2: Kondensasyonda oluşan asetoasetil-ACP’nin C-3’teki karbonil grubu indirgenir. Bu tepkime β-ketoaçil-ACP redüktaz ile katalizlenir ve elektron vericisi NADPH’tır.
-Basamak 3: Su, D-β-hidroksibutiril-ACP’nin C-2 ve C-3’ünden trans Δ2-butenoil-ACP ürününde çift bağ oluşturmak üzere uzaklaştırılır. Bu dehidrasyonu β-hidroksiaçil-ACP dehidrataz enzimi katalizler.
-Basamak 4: Çift bağın redüksiyonu: Son olarak D-β-hidroksibutiril-ACP’nin çift bağı, butiril-ACP oluşturmak üzere enoil-ACP redüktazla indirgenir, elektron vericisi yine NADPH’tır.
-Doymuş 4-C’lu yağ açili-ACP oluşumuyla yağ asidi sentaz kompleksinde bir tur tamamlanmış olur.
-Butiril grubu ACP’nin fosfopentatein-SH grubundan, β-ketoaçil-ACP sentazın, başlangıçtaki asetil grubunu taşıyan, Cys-SH grubuna taşınır.
-Zinciri iki karbon daha uzatacak dört tepkimeli bir sonraki döngüyü başlatmak için bir başka malonil grubu ACP’nin boş olan fosfopantetein-SH grubuna bağlanır.
-Kondensazyon ve redüksiyonun yedi turu, hala ACP’ye bağlı olan 16 C’lu doymuş palmitoil grubun oluşturur. Henüz anlaşılamamış nedenlerle zincir uzaması genellikle bu noktada durur.
-Serbest palmitat sentaz kompleksinin hidrolitik aktivitesi tarafından ACP molekülünden uzaklaştırılır.
-Bazı bitkilerde zincir sonlanması daha erken olur (8-14C uzunluğunda). Asetil CoA’dan palmitat sentezi için toplam tepkime iki kısımda düşünülebilir.birincisi yedi malonil CoA oluşumudur:
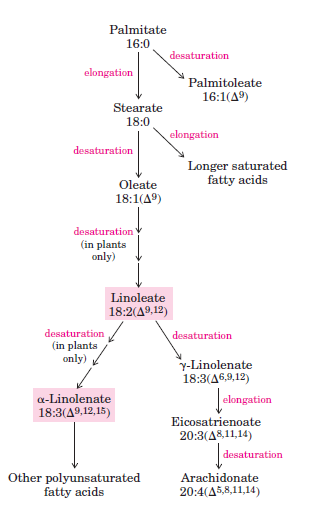
Uzun Zincirli Doymuş Yağ Asitleri Palmitattan Sentezlenir
-Mitokondri ve endoplazmik retikulumda bulunan yağ asiti uzatma sistemlerinin etkisiyle asetil gruplarının eklenmesi yoluyla uzatılabilir.
-Endoplazmik retikulumdaki uzatma mekanizması farklı enzim sistemlerinin olaya karışması ve tepkimede açil taşıyıcısının ACP’den daha ziyade koenzim A olması dışında palmitat sentezindekiyle aynıdır.
-Çift bağ, bir karışık işlevli oksidaz olan yağ açili-CoA desatüraz tarafından katalizlenen oksidatif bir tepkimeyle yağ asiti zincirine sokulur.
-İki farklı substrat olan yağ asidi ve NADPH, eş zamanlı olarak iki elektron oksidasyonuna uğrar.
-Memeli hepatositleri yağ asitlerinin Δ9 pozisyonuna kolaylıkla çift bağ sokar, ancak ek bir bağ sokamaz.
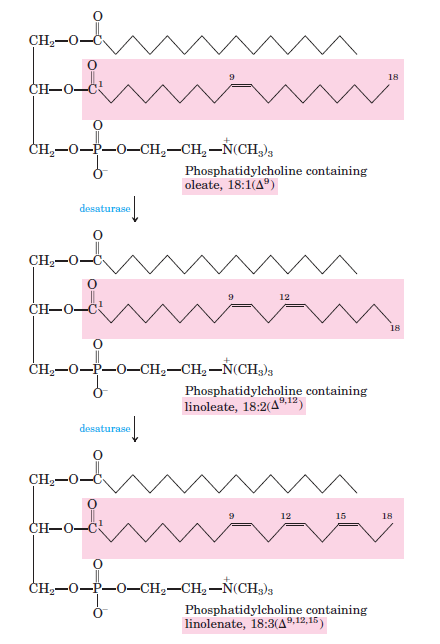
-Linoleik asit ve linolenik asitler ancak bitkiler tarafından sentezlenir ve esensiyel yağ asitleridir.
-Bitki desatürazları fosfotidilkolin-bağlı oleatı oksitleyerek, çoklu doymamış yağ asitlerini üretir. Ürünlerin bazıları fosfotidilkolinden hidrolizle ayrılır.








Biyokimya
-
Serum Enzimlerini Tayin Yöntemleri
-
Fosfatazlar (Alkali fosfataz= ALP)
-
Transferazlar
-
Transaminazlar
-
Enzimlerin Görev, İşlev ve Özellikleri - Enzimlerin İsimlendirilmesi
-
Kanda Bilirubin
-
Serum Proteinleri
-
Fosfolipidler
-
Trigliseridler
-
Kolesterol Nedir?
-
Kan Lipitleri Nelerdir?
-
Kan Şekeri Nedir?
-

Araşidonik Asit (ARA) Nedir?
-
Lizozim enzimleri
-
Lizozim: İlk Antibiyotik
