Ribozomların Biyosentezi
Tüm hücre komponentleri, ayrıcasız, biyosentez yoluyla, doğrudan yada dolaylı olarak DNA'dan köken alır. Ribozomların sentezi de DNA tarafından kodlanır. Ribozomların sentez yolu, çekirdekli hücrelerde (eukar-yotlarda) çekirdeksizlerden (prokaryotlardan) farklıdır.
Bacillussubtilis'de DNA'nın % OJ3'ü 16 S rRNA'yı ve % 0.25'i 23 S rRNA'yı kodlar. Bundan ve diğer organizmalarda yapılan gözlemlerden anlaşıldığı kadarıyla DNA'nın belirli bir kısmı (cistron) rRNA sfentezi için sorumludur.
Çekirdekli hücrelerde,ribozom sentezi, çekirdek içerisindeki çekirdekçiklerde yapılır. Çekirdekçikleri organize eden kfomozomal DNA bölgeleri, yani nukleolus-DNA'lar, rRNA kodlayan cistrona (operona) sahiptir. rRNA için nukleolar olmayan cistronların mevcut olup olmadığını bilemiyoruz. Eğer rRNA sentezinin zamansal süreci, örneğin sıçan beyninde incelenirce, aşağıdaki evreler görülür. Bu gözleme göre 18 rRNA (küçüğü), 28 S Rrna (büyüğü)'dan daha önce meydana gelmektedir. Sitoplazmada, tamamlanmış ribozomların meydana,gelmesi ise daha uzun bir zaman alır (genellikle 50 dakika civarında).
Bu 50 dakika içinde neler olduğunu kasaca görelim: rRNA'nın biyosentezi, bir seri öncül molekül dizisinden geçer, ilk oluşan 45 S'lik molekülden, farklı yol izlenerek, çıplak 18 S rRNA ve 28 S rRNA meydşr-,a gelir (Şekil 8.15/a). Tam bir ribozom ise daha sonra adım adım proteinlerle birleşmek suretiyle ortaya çıkar.
Hayvansal rRNA'ların biyosentezinde 45 S rRNA'dan itibaren birçok değişik
özellik görülür. tRNA'daki gibi, normal hazarın (A, G, U, C) yanı sıra, farklı nadir
bazları (pseudouridin, inosin ve özellikle adeninin, guaninin ve diğer bazların metil türevleri) içeren bu 45 Ş'lik ön madde, biyosentez sırasında, nadir bazların miktarı bakımından özgül değişiklikler gösterir. rRNA'nın olgunlaşması sırasında her adımda zincir biraz küçülür. Böylece G ve C'ce zehgin, pseudouridin ve metil gruplarınca fakir diziler elemine edilir.
5 S rRNA'ların sentezi için bağımsız bir cistronun (operofiun) varlığı gösterilmiştir. Bu cistronun büyüklüğü (sayıca), diğer rRNA türlerininkinin yaklaşık yarısı kadardır. 5 S rRNA'ların, büyük ribozomal alt birimler üzerine yığılımları çok geç olur. Bakteriyel 50 Ş ribozomların öncüsü olan 43 S partikülleri, 5 S rRNA taşımaz.
Ribozomal proteinlerin, rRNA'nın sentezini kodlayan cistronlar tarafından saptandığı savunulmaktadır. Bu durumda, 45 S rRNA ve bilinmeyen daha önceki moleküller, belirli bir zaman içerisinde mRNA işlevine sahiptir. 45 S'in öncü molekülünde bulunan kodonlar hesaplanırsa 46 x 105dalton/1000dalton = 4600 kodon bulunur.
Bu kodonların ise, tam bir ribozomda bulunan proteinlerin ancak %60’ını kodlamaya yeterli olacağı görülecektir. Buradan, rRNA’ların, ribozomal proteinlerin tümünü kodlayamamayacağı anlaşılır. Bunun ötesinde, çekirdekçilerde, ribozomal proteinlerin sentezi için herhangi bir bölgeye yada merkeze de rastlanmamıştır. Hem mikroorganizmalarda hem yüksek organizmaların hücrelerinde, ribozomların olgunlaşmasına bağımlı olmadan sentez edilen proteinlere de (kompertment proteinler) rastlanılmıştır.
Ribozomların biyosentezinde, birçok nokta açıklanamamıştır. Proteinlerin kökenleri, 45 S’lik öncül molekülden parçalanarak meydana gelen polinukleotitler ve 45 S rRNA’nın parçalanmasına katılan enzimler bilinmemektedir. rRNA’ların yapısına katılan nadir bazların işlevleri de tam olarak çözümlenememiştir.
Ribozomların tekrar tekrar kullanılması (turnover) fazla değildir. Yarılanma süreleri, karaciğer ribozomlarında yaklaşık 5 gün yada her saat için %0.4 ‘tür.
Ribozomların İşlevleri: mRNA’lar üzerinde bulunan bilgilerin, proteinlere aktarılmasını sağlarlar (translaşsyon) bunun için;
a) mRNA’lar üzerine özgül yığılımların olması ve bağlayıcı faktörlerin de katılması ile polizom oluşturularak, mRNA’nın 5 ucunun tanınması
b) Bilginin adım adım okunması,
c) Membran oluşumu ile translasyonun özelleşmesi sağlanır
Polizom Oluşumu: Yüz nukleotitten daha kısa olan polinukleotit dizileri, ribozomlara bağlanabildikleri halde, protein sentezi yapamazlar. En azından 150 nukleotitli diziler biyolojik olarak aktif birprotein sentezini gerçekleştirebilir. İnkubasyon ortamında, iyon değişiminin yükseltilmesi ile, kısa polinukleotit dizilerinin de protein sentezi yapması sağlanabilir.
mRNA’ların ribozomlara bağlanması, bağlayıcı bir faktörün katılması ile olur. Bu faktör hem mRNA’ların ribozomlara bağlanmasını, hem de başlama noktalarını yığışımların olmasını sağlar. Bu faktörler, gruplara özgü özellikler taşır ve ribozomlar üzerine ancak homolog mRNA’ların yığışımların oluşmasını sağlar. Bu faktörler, gruplara özgü özellikler taşır ve ribozomlar üzerine ancak homolog mRNA’ların yığışımını katalizler. Heterojen kaynaklardan gelenler normal koşullarda fiske olamaz.
Bununla beraber tek kollu DNA, dekstransülfat, polivvinilsülfat ve diğer mazı polianyonlar da ribozomlara bağlanabilir ve doğal polinukleotitlerle ribozomlar üzerinde rekabete girişirler.
mRNA ve ribozom arasındaki bağlanmaya polinukleotidin bazları katılmaz. Bağlanma, bir tarafta mRNA’nın fosfat (anyonik), diğer taraftan rRNA’nın, A,C ve G üzerindeki amino kökleri arasında olur. Bağlanma, ancak Ca, Mg yada Mn gibi iki değerli metal iyonlarının belirli bir derişiminde mümkündür. Derişimleri, 5 mM civarında olması gereken bu metaller, ribozomun arasında, mRNA’nın fiske edilmesine doğrudan doğruya katılmaz.
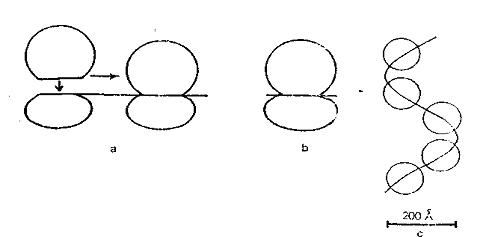
mRNA'ların bağlanması, Küçük ribozomal alt birimlerin aracılığıyla (30 S’lik veya 40 S'lik alt birimlerle) olur. Translasyon olayı ( ayrıntı proetin sentezinde) sırasında, küçük alt birimler ile büyük alt birimler karşı karsıya gelerek, ribozomları (70 S veya 80 S) meydana getirir. Öyleki, birçok ribozom, mRNA üzerinde tespit tanesi gibi dizilir. (Şekil 10). Öyleki, birçok ribozom, mRNA üzerinde tespit tanesi gibi dizilir. RICH, 1962 yılında, bu dizilmeye " Polizom " adını vermiştir. Daha önceki çalışmalarda tanımlanan serbest 70 S'li partüküller (tek tek ribozomlar) kısa mRNA parçası taşıyan yapısıdır. 70 S'li serbest ribozomların kökenleri ve işlevleri bilinmemektedir.
mRNA'lar ne kadar uzunsa, o kadar fazla ribozom taşıyacakları için farklı büyüklükte polizomlar ortaya çıkacaktır, iki ribozom arasındaki mesafe, yani bir ribozom tarafından okunan mRNA uzunluğu yaklaşık 90 nukleotite ulaşır. Bu ise 30 aminoasitlik kuramsal bir peplit zincirine denktir.
Özellikle büyük polizomlar, elektron mikroskobunda üç boyutlu tersiyer (üçüncül bir yapı (genellikle heliks) gösterir. Küçük alt birimler heliksin merkezine yönelmiştir. Polizomların büyüklüğü, dağılımı ve keza üçüncül yapısı, gelişim evrelerine özgül bir yapı gösterir. Embriyonik dokularda vida yada spiral şeklindeki polizomların ortaya çıkması, özel bir proteinin hücrede sentezlenmesiyle ilgili olduğu varsayılmaktadır.
Genetik
-

İnsanlarda Kaç Kromozom Vardır?
-
Sık görülen mikrodelesyon sendromları nelerdir?
-
Bilim insanları kromozomları nasıl inceler?
-
Arkea'da Kromozomlar ve DNA Replikasyonu
-
DNA Onarım Mekanizmaları Nelerdir?
-
DNA hasarına neden olan etkenler nelerdir?
-
XYY Süper Erkek Sendromu - JACOB’S, Sendromu
-
Bitki doku kültürü çalışmaları ile haploid bitkiler elde edilebilir
-
Gram pozitif bakterilerden genomik DNA izolasyon protokolü
-
E. coli bakterisinden genomik DNA izolasyon protokolü
-
DNA’nın Keşfi
-
İnsan Genom Projesi Nedir ? Amaçları Nelerdir ?
-
Genomik mikrodizilimlerle ikilenme teşhisi yöntemi
-
Gen duplikasyonu ve amplifikasyonu nedir?
-
DNA ile RNA Arasndaki Farklar ve Benzerlikler Nelerdir
